











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
淡水珍珠的生物矿化机理研究进展
引用本文
马玉菲, 乔莉, 冯庆玲. 淡水珍珠的生物矿化机理研究进展. 无机材料学报, 2013, 28(1): 109-116
MA Yu-Fei, QIAO Li, FENG Qing-Ling. Research Progress on Biomineralization Mechanism of Freshwater Pearl. Journal of Inorganic Materials, 2013, 28(1): 109-116
Permissions
MA Yu-Fei, QIAO Li, FENG Qing-Ling. Research Progress on Biomineralization Mechanism of Freshwater Pearl. Journal of Inorganic Materials, 2013, 28(1): 109-116
Copyright©2013, Editorial Board of Journal of Inorganic Materials
This is an open-access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
淡水珍珠的生物矿化机理研究进展
马玉菲(1987-), 女, 博士研究生. E-mail:mayf09@mails.tsinghua.edu.cn
摘要
碳酸钙广泛存在于生物矿物中, 是地球上最普遍的生物矿物之一。贝壳和珍珠的主要组成部分为碳酸钙无机相。我国淡水养殖珍珠多数品质优异, 具有良好的珍珠光泽。该种珍珠以文石晶型碳酸钙为无机相, 称为文石珍珠。近年来, 在我国淡水养殖珍珠中发现了球文石的存在, 球文石的出现是导致珍珠失去光泽、降低质量的主要原因。本文对比阐述了淡水文石珍珠和球文石珍珠的微观结构与性能, 总结了与珍珠层有关的体外模拟碳酸钙生物矿化的实验结果, 提出了珍珠层生物矿化机理未来的研究方向。
关键词:
淡水珍珠; 文石; 球文石; 生物矿化; 综述
中图分类号:P574
文献标志码:A
文章编号:1000-324X(2013)01-0109-08
Research Progress on Biomineralization Mechanism of Freshwater Pearl
Abstract
Calcium carbonate exists widely in many kinds of biominerals, and is one of the most widespread biominerals on the earth. The main component of the shell and pearl is calcium carbonate. In China, the majority of the freshwater pearls originated from
Keyword:
freshwater pearl; aragonite; vaterite; biomineralization; review
自然界的生物中存在着各种各样的生物矿物, 从细菌中的磁性体到贝壳、珊瑚、象牙、骨和牙齿, 即从微观纳米世界到宏观结构世界。在众多生物矿物中, 碳酸钙是存在范围最广、最普遍的生物矿物之一。大多数无脊椎动物中都发现了碳酸钙, 其中最具代表性的是珍珠和贝壳[ 1]。珍珠层作为珍珠和贝壳的主要结构, 是碳酸钙生物矿化研究的一个重要体系。我国浙江省诸暨淡水养殖三角帆蚌所培育的珍珠多数品质优异, 具有良好的珍珠光泽, 该种珍珠以文石晶型碳酸钙为无机相。近年来, 在我国淡水养殖珍珠中还发现了球文石的存在, 球文石晶体的出现是导致珍珠失去光泽、降低质量的主要原因。由于自然界中的球文石非常罕见, 目前国内外对于碳酸钙中的文石/方解石研究很多, 而对于文石/球文石的研究很少。因此, 文石和球文石珍珠的生物矿化是碳酸钙体系生物矿化机理研究的一个重要课题。然而, 存在于珍珠层中的球文石研究可以完善珍珠层以及碳酸钙的生物矿化理论体系。目前, 珍珠层生物矿化研究已经逐渐从认识生物矿物阶段发展到模仿生物矿物阶段, 这也在仿生学研究中具有重要意义。
1 文石珍珠和球文石珍珠结构与性能的对比研究
1.1 珍珠与珍珠层
珍珠是珍珠贝科动物马氏珍珠贝 Pteria martensii(Dunker)、蚌科动物三角帆蚌 Hyriopsis cumingii(Lea)或褶纹冠蚌 Cristaria plicata(Leach)等双壳纲软体动物的外套膜受到异物(沙粒、寄生虫)侵入的刺激, 受刺激处的表皮细胞以异物为核, 陷入外套膜的结缔组织中, 陷入的部分外套膜表皮细胞自行分裂形成珍珠囊, 珍珠囊细胞分泌珍珠质, 层复一层把核包被起来即可形成珍珠[ 2, 3]。珍珠层是构成珍珠和贝壳的主要结构。珍珠层的无机相为碳酸钙晶体, 约占总质量的95%, 由蛋白质-多糖构成的有机相约占5%。
1.2 珍珠层的生长机制
关于珍珠层的生长机制, 主要有以下四种理论: (1)细胞内部结晶、细胞外部组装; (2)隔室理论; (3)矿物桥理论; (4)模板理论。其中, 矿物桥和模板理论较完整地解释了珍珠层中堆垛和外延两种生长模式。
矿物桥理论认为, 通过不同珍珠层间有机质板片的孔隙, 文石晶体继续生长, 每个新形核的文石晶片沿套膜方向垂直生长, 直到碰到另一层层间基质, 垂直生长才会终止。随后, 板片横向生长形成新的板片。一旦正在生长的板片碰到板片上方相邻的层间基质中的孔隙时, 它将像矿物桥一样穿过孔隙使新的小板片继续结晶生长; 相对于下板片而言, 这个新板片存在横向偏移, 当较老的板片横向生长时, 在新老板片间形成更多矿物桥, 导致板片在较多位置上同时生长[ 1, 4, 5, 6]。
Weiner在研究贝壳有机质的生物化学组成时, 推断其可溶性有机质中存在高比例的(Asp-Y)2(Y: 表示Ser或Gly)序列, 如果蛋白质采用β折叠片结构, 则蛋白质中的Asp-Asp残基间距离(0.695 nm)正好与文石(或方解石)中的Ca2+-Ca2+ 距离(0.30~0.65 nm)相近, 因而认为可溶性有机质可能为矿物相结晶提供模板, 提出了珍珠层成因的模板理论[ 7, 8]。模板理论认为, 当无机相的某一晶面的结晶周期正好与带活性基团有机基质的结构周期相匹配时, 会降低无机矿物相晶体的形核活化能并诱导晶体沿此晶面方向生长, 从而导致晶体的有序定向结构。由于文石的(001)晶面的结晶学周期(即 a、 b)轴正好与有机基质的结构周期相匹配, 因而诱导文石晶体沿(001)晶面方向形核, 最终导致珍珠层中所有的文石晶片的 c轴垂直于珍珠层面; 此外, 当可溶性有机质在溶液中独立存在时, 同样由于晶格匹配而选择性地吸附于文石的(001)晶面上, 从而抑制文石晶体沿垂直该面方向生长, 致使文石晶体均形成板片状形貌。
Addadi等[ 9]经过总结研究提出了珍珠层沉积理论的基本观点: (1)类丝素纤维(silk-like)以凝胶状态存在, 预先填充在矿化区; (2)几丁质定向排列, 并控制碳酸钙晶体的定向生长; (3)首先形成的矿物相是胶体状的无定形碳酸钙; (4)形核在有机基质层面发生, 晶体在无定型碳酸钙(ACC)上生成; (5)酸性大分子在晶体生长的过程中起调控作用。珍珠层的生长包括4个主要过程: (1)有机基质的组装; (2)矿物相的初步形成; (3)单独文石板片形核; (4)文石板片生长形成成熟组织。
针对珍珠层的形成机制, 各理论均没有给出完善的结论, 通过对珍珠层不同方面的研究, 仅仅解释了各自观察到的一些现象。对珍珠层的研究仍在继续, 随着实验技术的不断改进, 珍珠层生长过程中的新现象必将引起关注。
1.3 文石珍珠和球文石珍珠的微观结构研究
通常, 我国淡水养殖的三角帆蚌所培育的珍珠比较圆润, 具有光泽, 大多数这种珍珠的无机相为文石晶型的碳酸钙。马红艳等[ 10]在研究浙江雷甸淡水珍珠时发现了球文石, 认为罕见的球文石取代文石是导致珍珠失去光泽的主要原因。因此, 这种干涩、瓷白色、无润感的珍珠也被称为球文石珍珠。文石珍珠和球文石珍珠的宏观光学照片如图1所示。
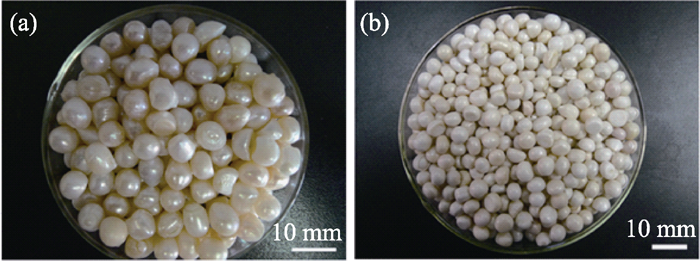 | 图1 文石珍珠(a)和球文石珍珠(b)的光学照片Fig. 1 Optical photos of aragonite pearls (a) and vaterite pearls (b) |
文石珍珠的珍珠层中碳酸钙晶体的形状、尺寸比较均匀, 通常为六边形, 晶片厚度约为0.25~ 0.99 μm, 长度约为3~5 μm, 每个板片外部均被有机基质包裹覆盖。文石板片层层堆积, 形成紧密的“砖墙结构”, 位于文石板片之间的有机层厚度约30 nm[ 11]。珍珠层中, 文石板片的 c 轴均与珍珠层面垂直[ 12, 13, 14, 15]。但关于珍珠层中不同文石板片 a、 b轴之间的关系仍未取得一致意见。Manne等[ 16]的研究表明, 双壳类( Atrinasp)珍珠层中不同板片的 a、 b轴在横向和垂向上均呈定向排列, 即整个珍珠层中所有晶体板片具有一致的结晶取向, 类似单晶。除此以外, 科研工作者还认为珍珠层中 a、 b轴的排列是生长过程中的内部竞争的结果[ 17]。Feng等[ 18]研究褶纹冠蚌( Cristaria plicata)的珍珠层时发现垂直于层面方向上相邻板片(3~10个) a、 b轴排列方向相同而构成取向畴结构。
Murr等[ 19]在研究中国淡水人工养殖三角帆蚌珍珠时发现该珍珠的内部(靠近珍珠核区)为方解石晶型碳酸钙, 外部(靠近珍珠表面)为文石晶型碳酸钙。该珍珠内、外部的微观结构如图2所示。
 | 图2 珍珠内层晶体(方解石)(a)及外层晶体(文石)(b)的扫描电镜照片[ 19]Fig. 2 Comparative SEM views for pearl internal layer crystals (mostly calcite) (a) and external surface crystals (mostly aragonite) (b)[ 19] |
Qiao等[ 20]研究了球文石珍珠层的结构和形貌(图3), 认为球文石板片的结构与文石板片的结构类似。截面(长×宽)为平行于珍珠表面方向的堆垛方式, 板片平均尺寸为8 μm×2 μm。截面(长×高)为典型的砖墙型堆垛结构, 球文石板片厚度小于正常的文石板片, 且显得疏松。截面(宽×高)形状为斜四边形, 尺寸约为2.0 μm×0.4 μm。这种形貌支持矿物桥理论解释的“堆垛生长”, 也就是说, 在不同层相邻的板片轴心在同一直线上, 这种区域可以跨越数层距离。此外, 张刚生等[ 21]在研究诸暨三角帆蚌养殖珍珠时, 发现珍珠表面层是由两种完全不同形貌的球文石组成的。第一, 球文石晶体呈不规则六边形薄片状, 晶体的长径约为4.4~7.1 μm, 短径约为1.4~2.6 μm; 第二, 晶体呈纤维柱状, 每个纤维柱由更细的纤维聚集形成, 纤维柱的长径为7.0~11.6 μm, 短径为0.9~1.5 μm。
 | 图3 球文石珍珠层的三维微观结构和示意图[ 20]Fig. 3 SEM images of vaterite tablets and layers in lackluster pearls after 10wt% EDTA-2Na treatment[ 20](a) Etched surface of lackluster pearl (scale of amplificatory image, 1 μm); (b) Etched section of length by thickness of vaterite tablets; (c) Etched section of width by thickness of vaterite tablets; (d) Three- dimensional structure sketch of vateite crystal in lackluster pearls. |
纳米压痕技术可以研究文石和球文石珍珠抛光截面的力学性能。通过测定两种珍珠截面的平均硬度随压头深度的变化, 发现文石珍珠截面的平均硬度为3.032 GPa, 球文石珍珠截面的平均硬度为1.301 GPa; 文石珍珠的硬度是球文石珍珠的2.7倍[ 22]。矿物的硬度与晶体结构中化学键型﹑原子间距﹑电价和原子配位等密切相关。此外, 球文石珍珠和文石珍珠的硬度还与珍珠本身的微结构有关。方解石晶体的硬度比文石晶体低, 但在贝壳外部的棱柱层是由柱状方解石单元紧密结合而成, 其硬度反而比内部的珍珠层高, 可以起到保护和防御作用。而对于球文石来说, 无论是天然球文石, 人工球文石还是生物体内的球文石颗粒, 球文石都难以形成致密的结构, 其疏松的微观结构很可能是导致球文石珍珠硬度降低的主要原因。
文石珍珠层具有晶体学择优取向。Qiao等[ 22]利用微区X射线衍射法对球文石表面800 μm× 800 μm的区域进行了晶体学取向表征(图4)。结果表明球文石板片同样存在较强的择优取向, 其织构方向为[102]和[101], 即[010]晶向存在较强的择优取向, 且[010]晶向垂直于球文石板片表面[ 23]。该研究证实了球文石板片构成的微观结构和文石珍珠层一样, 存在强的择优取向, 球文石晶体 b轴沿珍珠层表面的法线方向。
文献报道, 文石珍珠层内部存在大量孪晶, 这与文石的晶体结构有直接关系, 无论是地质文石还是生物文石, 都很容易产生孪晶。Qiao等[ 23]利用高分辨透射电子显微镜研究了球文石珍珠层的内部结构(图5(a))。区域A是典型的孪晶堆垛层错, 区域B出现了调制结构。在与该区域对应的选区电子衍射结果(图5(b))中也发现了(001)层错面的堆垛层错结构和调制结构。
1.4 珍珠中文石和球文石的稳定性研究
碳酸钙有三种主要晶型: 方解石、文石和球文石, 其中, 球文石是最不稳定的。球文石在去离子水中20 h可完全转变为方解石[ 24]。但是, 在珍珠体系中, 球文石的稳定性和文石晶体基本相同, 是一种稳定相。自然界中的球文石在较低温度下就能转变成方解石, 而淡水珍珠中的球文石晶体其相变温度与文石相变温度基本相同, 均为520℃[ 25]。因此, 可以认为珍珠中球文石晶体的热稳定性与文石晶体基本相同。
自然界中, 球文石在水溶液中很快被溶解, 溶解度很高。为了研究两种珍珠在溶液中的稳定性, Qiao等[ 25]选取去离子水、乙二胺四乙酸钠(Na2EDTA)溶液及柠檬酸溶液, 分别代表中性水溶解、络合溶解和有机酸溶解三种腐蚀碳酸钙的方式。研究发现, 球文石粉末可以在去离子水中浸泡20 d而不发生相变; 在Na2EDTA和柠檬酸的溶液中, 球文石和文石的腐蚀溶解速度基本一样。所以, 球文石珍珠在水溶液中具有与文石珍珠类似的稳定性, 这也是球文石珍珠可以和文石珍珠长期共存的主要原因。
球文石稳定存在的原因, 主要有以下几点: (1)有机基质的作用。球文石稳定存在于生命活动中, 主要是因为有机基质可以稳定球文石的晶型和结构, 以前的研究证明, 从含有球文石晶体的生物矿物中提取的有机基质可以直接诱导球文石晶体生长[ 26]。(2)复杂的微观结构。生物矿物一般均具有复杂的分级结构和纳米结构, 复杂的结构以及纳米尺度的结构会发生动力学抑制, 使得原本不稳定的结构稳定存在[ 27, 28]。(3)板片内部存在缺陷。珍珠层板片内部存在大量的孪晶和堆垛, 堆垛层错作为晶体的生长台阶, 诱发晶体在相对低的能量下生长。
2 体外模拟碳酸钙晶体生长研究
2.1 珍珠层中的生物大分子作为添加剂的体外矿化研究
珍珠层中的有机基质是以柔韧的有机片层的形式存在的。这种有机片层均为复合物, 由三种生物大分子组成: (1) 不可溶的几丁质, 呈β折叠片结构, 位于有机基质的中心; (2) 富含甘氨酸和丙氨酸的不溶性蛋白, 具有反平行β折叠片结构, X射线衍射发现其结构和组成与丝纤维类似, 因此被称为丝蛋白, 位于几丁质的上、下表面; (3) 富含天冬氨酸等酸性氨基酸的可溶性蛋白, 具有β折叠片结构, 直接与珍珠层接触。在珍珠层的生物矿化过程中, 酸性有机质对文石晶体的形成起重要作用, 有机质中的酸性侧链与钙离子之间具有强烈的亲合作用, 从而形成文石晶体的结晶中心[ 29]。
最早开始研究珍珠层中有机基质生物化学的是Creenshaw[ 30], 他采用弱酸脱除碳酸钙矿物后获得了珍珠层中较完整的生物大分子。此后, 有关珍珠层中有机基质的研究揭示了一些普遍规律: 珍珠层中的有机基质主要为蛋白质, 分为可溶性基质(SM)和不溶性基质(IM), SM一般为酸性糖蛋白, 亲水; IM为偏碱性糖蛋白, 疏水。
体外模拟矿化是研究生物矿物中有机基质功能的主要方法。James等[ 31]利用EDTA提取出鲍鱼壳珍珠层的可溶基质。研究发现, 在过饱和碳酸钙溶液中引入这种有机质, 方解石籽晶上可生长文石晶体, 碳酸钙(001)面直接生长在方解石的(1014)面。另外, Feng[ 32]、Belcher[ 33]、Falini[ 34]、Samata[ 35]、Kono[ 36]等在对其他碳酸钙生物矿物的研究中发现, 提取出的可溶性有机质可以在体外实验中诱导不同晶型的碳酸钙, 而不溶性有机质在晶体生长中起模板作用。
Ma等[ 37]采用水和弱酸从淡水文石珍珠和球文石珍珠中分别提取了水可溶有机质(WSM)、酸可溶有机质(ASM)及酸不溶有机质(AIM)并进行碳酸钙晶体的体外模拟矿化研究。发现文石珍珠的水溶有机质能诱导文石晶体生长, 球文石珍珠的酸溶有机质能诱导生长球文石晶型的碳酸钙(图6).
 | 图6 文石珍珠水可溶有机质(a)及球文石珍珠酸可溶有机质(b)诱导的碳酸钙晶体[ 37]Fig. 6 Calcium carbonate crystals induced by WSM of aragonite pearl (a) and ASM of vaterite pearl (b)[ 37] |
Ma等[ 38]以文石珍珠的水可溶有机质作为添加剂, 研究了不同矿化微环境下有机质对碳酸钙晶体的调控作用。当矿化液pH变化时, 文石晶体主要受WSM调控。矿化液pH值对方解石晶体影响显著。未添加WSM, 随pH增大, 方解石逐渐形成不规则形貌。添加WSM, 方解石随pH增大逐渐恢复菱面体形貌。矿化液温度能够显著影响碳酸钙晶体的晶型和形貌。当矿化液温度在2~70℃范围变化时, 碳酸钙晶体由球文石向文石转变。在此过程中, WSM对碳酸钙晶体生长无显著调控作用。
Hou等[ 39]利用从贝壳珍珠层中提取的不溶蛋白膜作为模板进行碳酸钙的体外模拟矿化实验。当只有蛋白膜存在时, 合成的碳酸钙晶体为规则的菱面体形貌, 实验结果证实了棱柱层的不溶性基质不具有诱导形核的作用, 但当甘氨酸或天冬氨酸与蛋白膜同时存在时, 方解石出现了菠萝状聚集体形貌。
碳酸钙体外模拟矿化实验表明, 从含有文石晶体的生物矿物中提取出来的可溶基质可以诱导文石晶型碳酸钙。Belcher等[ 33]从鲍鱼 Haliotis refescens壳层中提取的可溶有机质显示了很好的对应性, 该结果显示可溶性基质蛋白能单独控制CaCO3的晶型和形貌, 不溶性基质蛋白并不是必需的。Feng等[ 32]从贝壳中提取的水可溶有机质也可以控制文石晶体的生长, 而不溶性有机质只是影响晶体的尺寸和密度。
2.2 珍珠层和氨基酸层为基底的体外矿化研究
对于珍珠层中文石板片形成的矿化机理, 目前存在以下三种假设: (1)单晶生长, 单晶片可以很好地结合传统的矿物桥或异相外延生长理论; (2)纳米晶粒聚合体; (3)不稳定的无定形碳酸钙或球文石相转变。Nassif等[ 40]在 Haliotis laevigate珍珠层中发现一层连续的无定形碳酸钙层, 该无定形碳酸钙是相变过程中残留的产物; Rousseau等[ 41]发现 Pinctada maxima珍珠层中的单个文石板片是由有机基质包裹的纳米晶粒组成。因此, 对于珍珠层单个板片的形成过程, 还没有统一的理论。
Qiao等[ 42]利用新鲜的文石珍珠层、球文石珍珠层和氨基酸改性的方解石表面作为模板, 在过饱和碳酸钙溶液中进行体外模拟矿化来研究珍珠层的生长机理。当新鲜的文石珍珠断面作为模板时, 可生长出一层文石“类珍珠层板片”(图7)。生长过程依次经历了无定形碳酸钙、定向的纳米文石堆垛结构和不规则的六边形板片。这些不规则的六边形文石晶体具有和天然珍珠层中的文石板片相似的形貌和结构, 且 c轴垂直于层面。生长过程中, 新鲜珍珠表面的有机基质可吸附钙离子和碳酸根离子, 无定形碳酸钙可能作为前驱相存在。图8为珍珠层上碳酸钙晶体生长的示意图。当新鲜的球文石珍珠层作为矿化模板时, 可以直接诱导生成稳定的球文石晶体(图9), 体现了球文石珍珠有机基质对碳酸钙晶型的调控作用。当谷氨酸改性的方解石表面作为模板时, 可生长出定向的方解石层(图10(a)~(c))。如果在新生长的方解石表面重复谷氨酸沉积和晶体生长的过程, 可以得到具有一致的晶体学取向的双层方解石层(图10(d))。氨基酸分子被认为在传递两层方解石之间的取向信息上起到重要作用[ 43]。
 | 图7 “类珍珠层板片”的生长过程[ 42]Fig. 7 SEM images of CaCO3 grown on nacre surface for different time[ 42](a) 10 min; (b) 30 min; (c) 1 h; (d) 2 h; (e) 3 h; (f) 10 h |
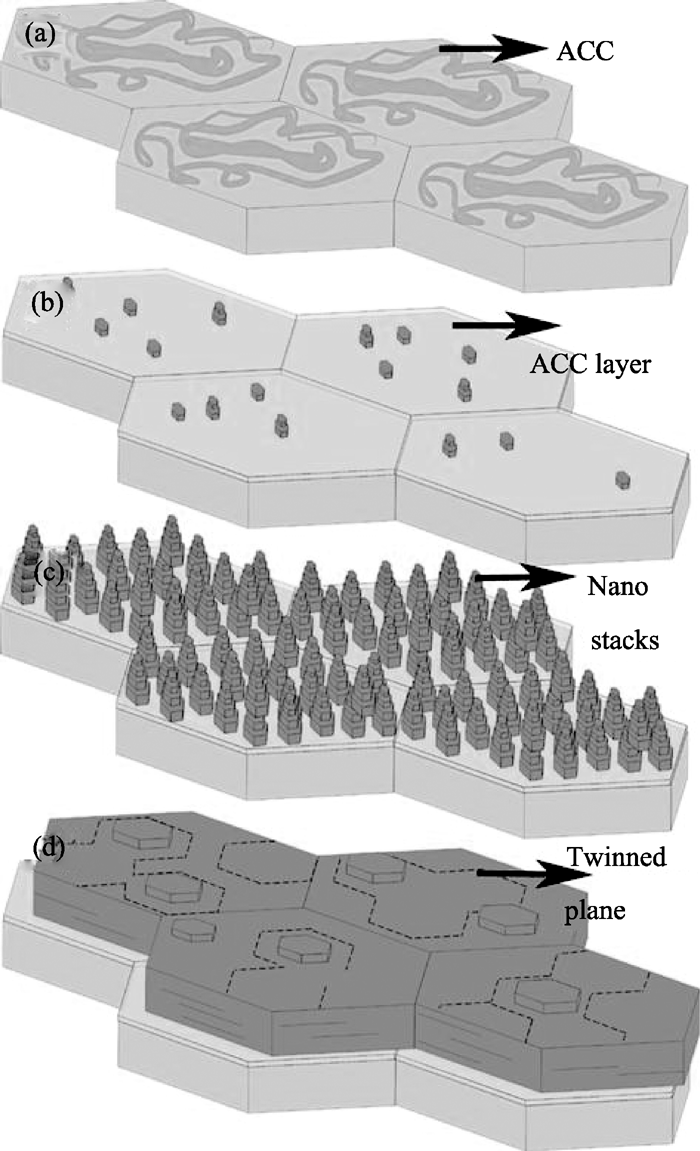 | 图8 原始珍珠层上碳酸钙晶体生长模型图[ 42]Fig. 8 Schematic representation of the growth sequence of CaCO3 on nacre[ 42](a) ACC; (b) ACC layer; (c) Nano stacks; (d) Twinned plane |
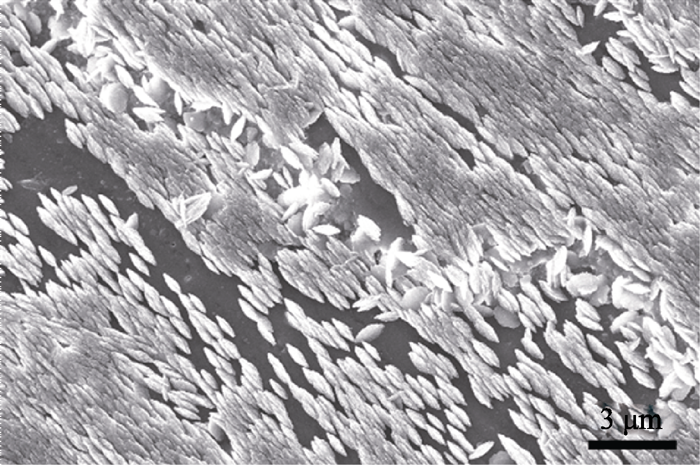 | 图9 在球文石珍珠层表面生长的球文石晶体的SEM照片[ 22]Fig. 9 SEM image of vaterite crystals grown on the original section of vaterite pearl[ 22] |
 | 图10 谷氨酸改性的方解石基底上生长的方解石晶体[ 43]Fig. 10 SEM micrographs of calcite matrix and calcite layers deposited on glutamic acid layers[ 43](a) Calcite tablets grown for 1 h; (b) the bigger and conjunctive calcite tablets grown for 1.5 h; (c) the calcite layer grown for 2 h; (d) SEM micrograph of double layered structure of calcite and glutamic acid |
3 总结与展望
由于珍珠层生物矿化本身的复杂性, 难以在原位进行生长研究, 大多利用各种模型对其进行基础性研究, 目前的研究重点在于明确生物矿化的分子机理。另一方面, 有关珍珠层生物矿化的相关生物学问题仍不清楚, 在生物矿化中生物控制的一些基本问题仍处于探索阶段。关于生物矿化过程中细胞、基质、矿物质三者间的互动关系, 是生物矿化调控的基础[ 1]。遗憾的是, 我们对其中很多关键问题尚缺乏清楚的认识。虽然过去几十年珍珠层生物矿化的研究已经取得了一些成就, 特别是关于有机基质调控矿化的机理方面取得了共识。但是目前对于生物矿化过程的认识还处于较低的水平, 仍有许多问题尚待解决, 如: 如何模拟生物微环境进行生物矿化研究, 如何对珍珠层生物矿化过程进行实时监控, 如何建立珍珠层生物矿化过程中细胞、基质、矿物质间的互动关系模型等。因此, 未来珍珠层生物矿化研究要提高到一个新的水平, 即把生物体内的矿化当作生命体系中的一个子系统来进行研究。对于珍珠层来说, 将软体动物生命过程中的基本特征与珍珠及贝壳的形成联系起来, 软体动物受刺激后如何启动、控制病理矿化过程形成珍珠, 即珍珠囊上皮细胞分泌有机基质和钙离子, 精确调控珍珠层的自组装过程; 以及正常的生理矿化过程形成的贝壳珍珠层。随着研究的逐渐深入, 从蛋白调控进入细胞、基因调控研究, 此类研究必将产生更多的具有潜在应用价值的新理论和新方法。
参考文献
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
|
| [31] |
|
| [32] |
|
| [33] |
|
| [34] |
|
| [35] |
|
| [36] |
|
| [37] |
|
| [38] |
|
| [39] |
|
| [40] |
|
| [41] |
|
| [42] |
|
| [43] |
|